2° Ano E Biologia Profa Solange
Período para entrega: Até 16/09/2020
Unidade: DNA como material genético- DNA- Receita da Vida e seu Código.
Instruções:
1- Fazer a Leitura e Interpretação de Texto- (Khan Academy)- Texto 1- Descoberta da estrutura do DNA
Texto 2- Experimentos Clássicos: DNA como material genético
2- Responder as questões disponibilizadas abaixo dos textos.
Descoberta da estrutura do DNA
A estrutura de dupla hélice do DNA e como foi descoberta. Chargaff, Watson e Crick, e Wilkins e Franklin.
Introdução
Hoje, a dupla hélice de DNA é provavelmente a mais icônica de todas as moléculas biológicas. Inspirou escadas, decorações, pontes de pedestres (como a de Singapura, mostrada abaixo), e mais.
Tenho que concordar com os arquitetos e designers: a dupla hélice é uma estrutura linda, e sua forma se encaixa com sua função de forma notável. Mas a dupla hélice nem sempre foi parte do léxico cultural. Na verdade, até meados da década de 1950, a estrutura do DNA era um mistério.
Neste artigo, vamos brevemente explorar como a estrutura de dupla hélice do DNA foi descoberta através do trabalho de James Watson, Francis Crick, Rosalind Franklin, e outros pesquisadores. Então, veremos as propriedades da dupla hélice.
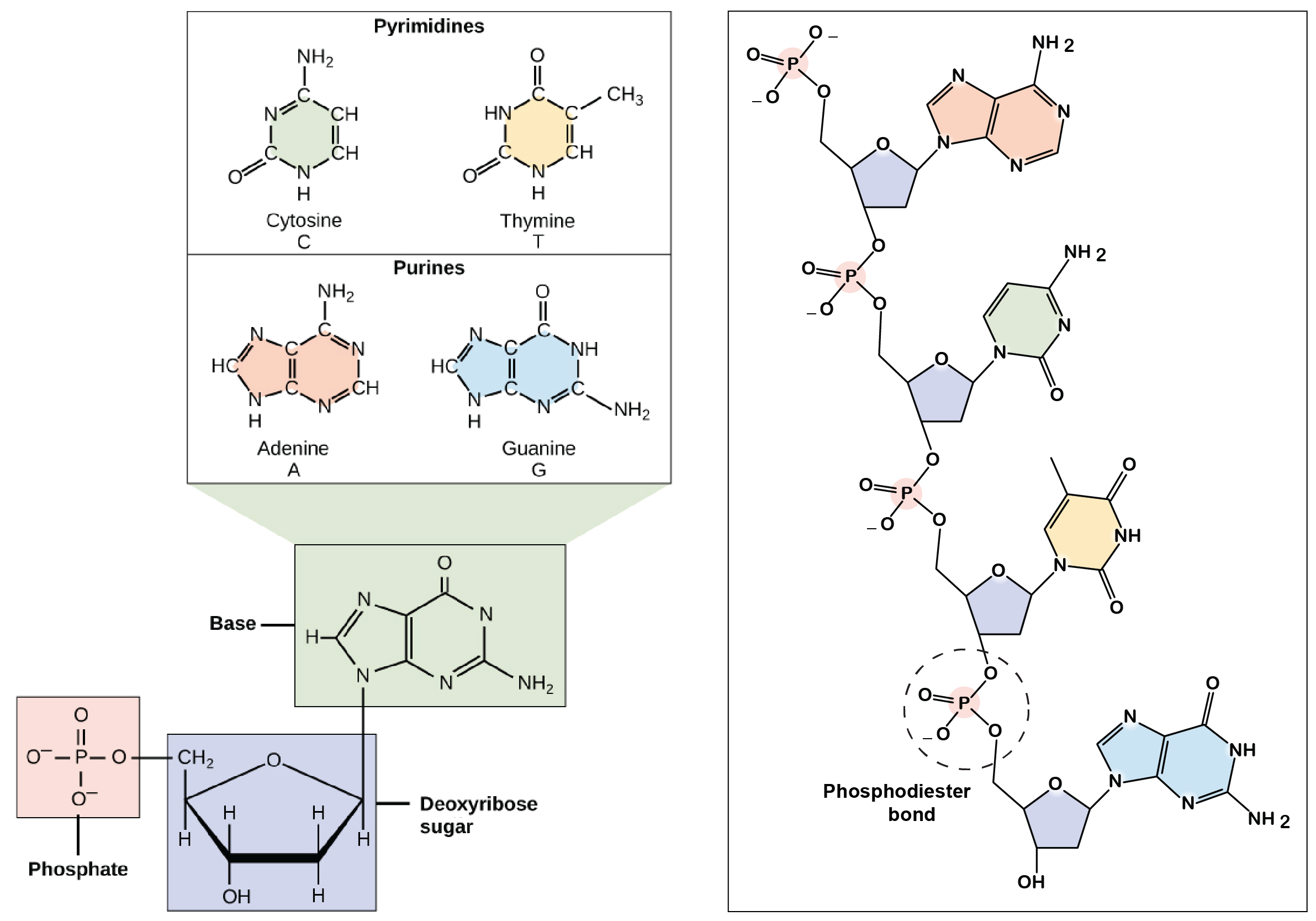
Os componentes do DNA
Por conta do trabalho do bioquímico Phoebus Levene e outros, os cientistas na época de Watson e Crick sabiam que o DNA era composto de subunidades chamadas
nucleotídeos1start superscript, 1, end superscript. Um nucleotídeo é feito de um açúcar (desoxirribose), um grupo fosfato, e uma das quatro bases nitrogenadas: adenina (A), timina (T), guanina (G) ou citosina (C).
As bases C e T, que têm apenas um anel, são chamadas de piridiminas, enquanto as bases A e G, que têm dois aneis, são chamadas de purinas.
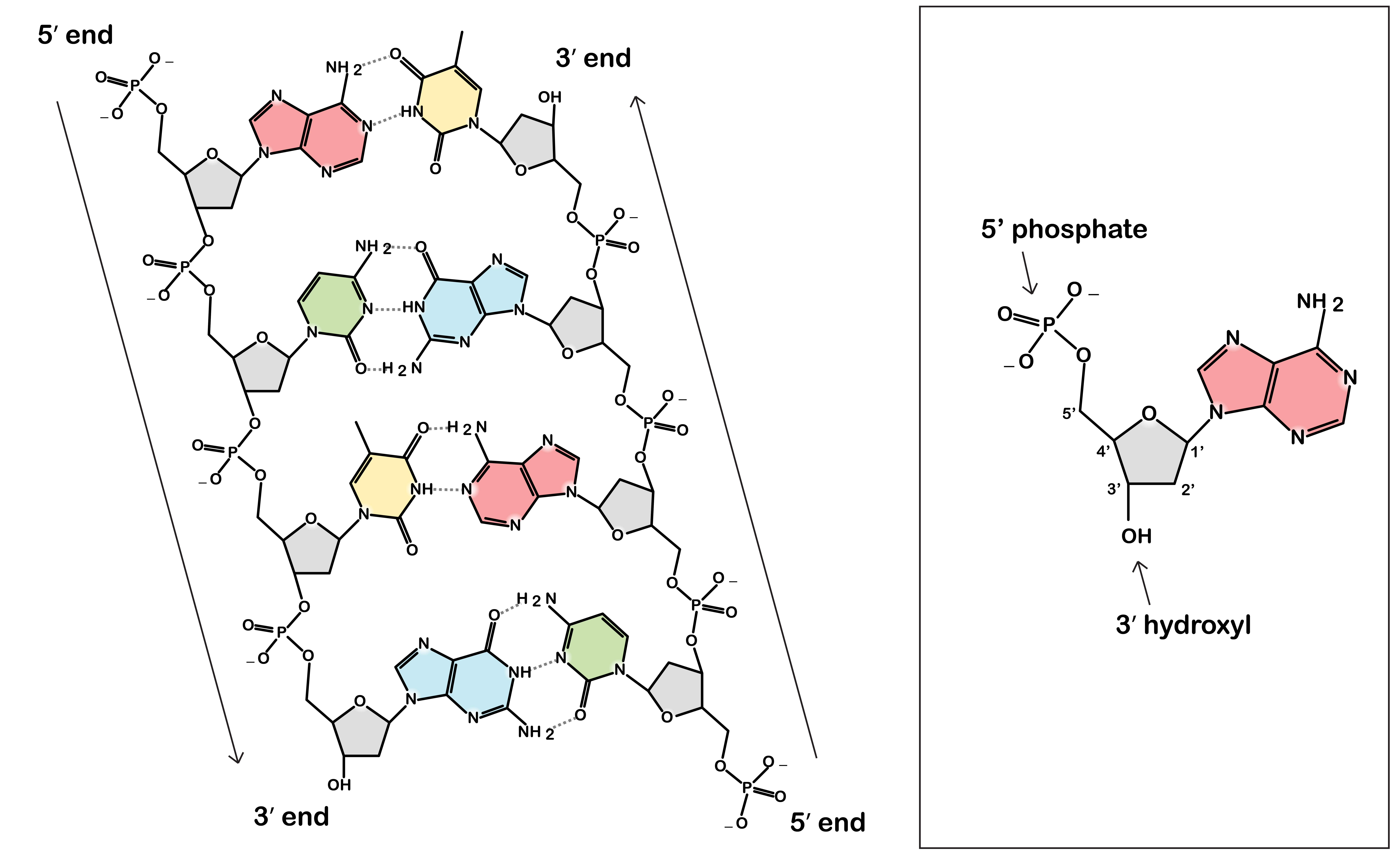
Os nucleotídeos de DNA se reúnem em cadeias ligadas por ligações covalentes, que se formam entre o açúcar desoxirribose de um nucleotídeo e o grupo fosfato do próximo. Esse arranjo faz uma cadeia alternada de grupos fosfato e de açúcar desoxirribose no polímero DNA, uma estrutura conhecida como esqueleto de açúcar-fosfato
Regras de Chargaff
Outra informação chave relacionada à estrutura de DNA veio do bioquímico austríaco Erwin Chagaff. Chargaff analisou o DNA de diferentes espécies, determinando sua composição de bases A, T, C e G. Ele fez várias observações fundamentais:
A, T, C e G não eram encontradas em quantidades iguais (como alguns modelos da época diziam)
As quantidades de bases variavam entre as espécies, mas não entre indivíduos da mesma espécie
A quantidade de A sempre igualava a quantidade de T, e a quantidade de C sempre igualava a quantidade de G (A = T e G = C)
Esses resultados, chamados de regras de Chargaff, acabaram sendo cruciais para o modelo de Watson e Crick da dupla hélice de DNA.
Watson, Crick e Rosalind Franklin
No início dos anos de 1950, o biólogo americano James Watson e o físico britânico Francis Crick elaboraram seu famoso modelo da dupla hélice de DNA. Eles foram os primeiros a cruzar a linha de chegada nessa "corrida" científica, com outros como Linus Pauling (que descobriu a estrutura secundária da proteína) também tentando encontrar o modelo correto.
Em vez de fazer novos experimentos no laboratório, Watson e Crick coletaram e analisaram conjuntos de dados já existentes, organizando-os de formas novas e esclarecedoras
2squared. Algumas de suas pistas mais cruciais sobre a estrutura do DNA vieram de Rosalind Franklin, uma química que trabalhava no laboratório do físico Maurice Wilkins.
Franklin era uma especialista em uma técnica poderosa para determinar a estrutura das moléculas, conhecida como cristalografia de raios-x. Quando a forma cristalizada de uma molécula como o DNA é exposta a raios-x, alguns dos raios são defletidos pelos átomos no cristal, formando um padrão de difração que dá pistas sobre a estrutura da molécula.
A cristalografia de Franklin deu a Watson e Crick pistas importantes para a estrutura do DNA. Algumas vieram da famosa "imagem 51", uma imagem de raio-x de difração do DNA notavelmente clara e impressionante produzida por Franklin e seus estudantes de graduação (um exemplo moderno do padrão de difração produzido pelo DNA é mostrado abaixo). Para Watson, o padrão de difração em formato de X da imagem de Franklin imediatamente sugeriu uma estrutura helicoidal, de duas fitas para o DNA
3cubed.
Watson e Crick juntaram dados de um número de pesquisadores (incluindo Franklin, Wilkins, Chargaff e outros) para montar seu celebrado modelo da estrutura de DNA em 3D. Em 1962, James Watson, Francis Crick e Maurice Wilkins foram premiados com o prêmio Nobel de medicina. Infelizmente, a essa altura Franklin já havia morrido, e os prêmios Nobel não são concedidos a título póstumo.
O modelo do DNA de Watson e Crick
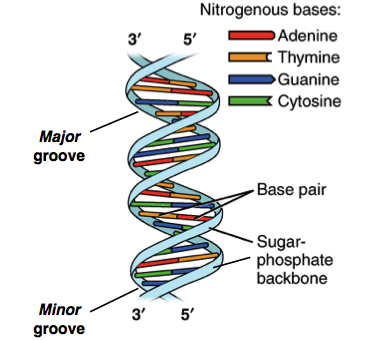
A estrutura do DNA, como representada no modelo de Watson e Crick, é uma hélice de dupla fita, antiparalela, para a direita. Os esqueletos de açúcar-fosfato das fitas de DNA constituem o exterior da hélice, enquanto as bases nitrogenadas são encontradas no interior e formam pares ligados por ligações de hidrogênio que mantêm as fitas de DNA juntas.
No modelo abaixo, os átomos laranja e vermelho marcam os fosfatos dos esqueletos de açúcar-fosfato, enquanto os átomos azuis no interior da hélice pertencem às bases nitrogenadas.
Orientação antiparalela
O DNA de fita dupla é uma molécula antiparalela, ou seja, é composta de duas fitas que correm lado a lado mas apontam para direções opostas. Em uma molécula de DNA de fita dupla, a extremidade 5' (com fostato livre) de uma fita se alinha com a extremidade 3' (com hidroxila livre) de sua parceira, e vice-versa.
Hélice dextrógira
No modelo de Watson e Crick, as duas fitas de DNA enrolam-se uma em volta da outra para formar uma
hélice dextrógira. Todas as hélices têm uma direção, que é uma propriedade que descreve como seus filamentos são orientados no espaço.
A torção da dupla fita de DNA e a geometria das bases criam um vão maior (chamado de sulco maior) e um vão menor (chamado de sulco menor) que estão ao longo do comprimento da molécula, como mostrado na figura acima. Esses sulcos são importantes locais de ligação para proteínas que mantêm o DNA e regulam a atividade dos genes.
Pareamento de bases
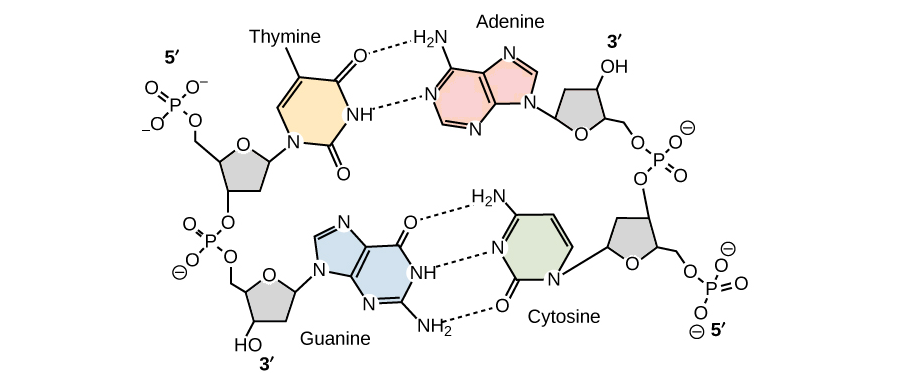
No modelo de Watson e Crick, as duas fitas da dupla hélice de DNA são mantidas juntas por ligações de hidrogênio entre as bases nitrogenadas nas fitas opostas. Cada par de bases fica plano, formando um "degrau" da escada da molécula de DNA.
Pares de base não são feitos de qualquer combinação de bases. Em vez disso, se há um A em uma fita, ele deve ser pareado com um T na outra (e vice-versa). Similarmente, um G encontrado em uma fita, deve sempre ter um C como parceiro na fita oposta. Essas associações A-T e G-C são conhecidas como pares de base complementares.
O pareamento de bases explica as regras de Chargaff, ou seja, porque a composição de A é sempre igual a de T, e a composição de C se iguala a de GG
11start superscript, 11, end superscript. Onde há um A em uma fita, deve haver um T na outra, e o mesmo é verdade para G e C. Porque uma grande purina (A ou G) é sempre pareada com uma pequena piridimina (T ou C), o diâmetro da hélice é uniforme, chegando a cerca de
22 nanômetros.
Apesar do modelo original de Watson e Crick propor que haveriam duas ligações de hidrogênio entre as bases de cada par, sabemos hoje que G e C formam uma ligação adicional (de forma que os pares A-T formam duas ligações de hidrogênio, enquanto pares G-C formam três)
12start superscript, 12, end superscript.
O impacto da dupla hélice
A estrutura do DNA abriu a porta para a compreensão de muitos aspectos da função do DNA, como saber como ele é copiado e como a informação que carrega é usada pela célula para fazer proteínas.
Assim como veremos nos próximos artigos e vídeos, o modelo de Watson e Crick inaugurou uma nova era de descobertas em biologia molecular. O modelo e as descobertas que ele permitiu formam os fundamentos da maioria das pesquisas de ponta em biologia e biomedicina atualmente.
Explore além da Khan Academy
Você quer aprender mais sobra a escada do DNA? Confira esta
atividade interativa (em inglês) do LabXchange.
LabXchange é uma plataforma on-line gratuita de educação científica criada na Faculdade de Artes e Ciências de Harvard e apoiada pela Fundação Amgen.
Créditos:
Este artigo foi produzido com base nos seguintes artigos:
Referências:
B-form, A-form, and Z-form of DNA. (2014, May 4). Acesso em 27 de julho, 2016. Disponível em bio.libretexts.org/Core/Genetics/Unit_I%3A_Genes,_Nucleic_Acids,_Genomes_and_Chromosomes/2%3A_Structures_of_nucleic_acids/B-Form,_A-Form,_Z-Form_of_DNA.
Referências:
B-form, A-form, and Z-form of DNA. (2014, May 4). Acesso em 27 de julho, 2016. Disponível em bio.libretexts.org/Core/Genetics/Unit_I%3A_Genes,_Nucleic_Acids,_Genomes_and_Chromosomes/2%3A_Structures_of_nucleic_acids/B-Form,_A-Form,_Z-Form_of_DNA.
Purves, W. K., Sadava, D. E., Orians, G. H., and Heller, H.C. (2004). The structure of DNA. In Life: The science of biology (7th ed., pp. 217-220). Sunderland, MA: Sinauer Associates.
Reece, J. B., Urry, L. A., Cain, M. L., Wasserman, S. A., Minorsky, P. V., and Jackson, R. B. (2011). Building a structural molecule of DNA: Scientific inquiry. In Campbell biology (10th ed., pp. 316-318). San Francisco, CA: Pearson.
Experimentos Clássicos: DNA como material genético
Experimentos por Frederick Griffith, Oswald Avery e seus colegas, e Alfred Hershey e Martha Chase.
Introdução
Nosso entendimento do papel do DNA na hereditariedade nos leva a uma variedade de aplicações práticas, incluindo análises forenses, testes de paternidade e mapeamento genético. Graças a essas várias utilidades, atualmente muitas pessoas conhecem o conceito de DNA.
Pode até parecer surpreendente, mas menos de um século atrás, nem mesmo os membros da comunidade científica mais bem educados sabiam que o DNA era nosso material genético.
Neste artigo examinaremos alguns experimentos clássicos que levaram à identificação do DNA como o portador da informação genética.
Proteína x DNA
O trabalho de
Gregor Mendel mostrou que características (como a coloração da flor em plantas de ervilha) não eram herdadas diretamente, mas ao invés, eram especificadas por genes que passam dos pais para a prole. O trabalho de outros cientistas por volta do século 20, incluindo Theodor Boveri, Walter Sutton e
Thomas Hunt Morgan, estabeleceram que os fatores hereditários de Mendel eram, provavelmente, transmitidos pelos cromossomos.
Cientistas inicialmente pensaram que as proteínas, que são encontradas em cromossomos ao longo do DNA, seriam as componentes do tão procurado material genético. Proteínas eram conhecidas por suas sequências diversas de aminoácidos, enquanto se pensava que o DNA era um polímero entediante e repetitivo, devido, em parte, a um modelo incorreto (mas popular) de sua estrutura e composição
1start superscript, 1, end superscript.
Hoje, sabemos que o DNA na verdade não é repetitivo e pode carregar grandes quantidades de informação, como discutido mais profundamente no artigo sobre a
descoberta da estrutura do DNA. Mas como os cientistas inicialmente perceberam que o "entediante" DNA talvez pudesse ser o material genético?
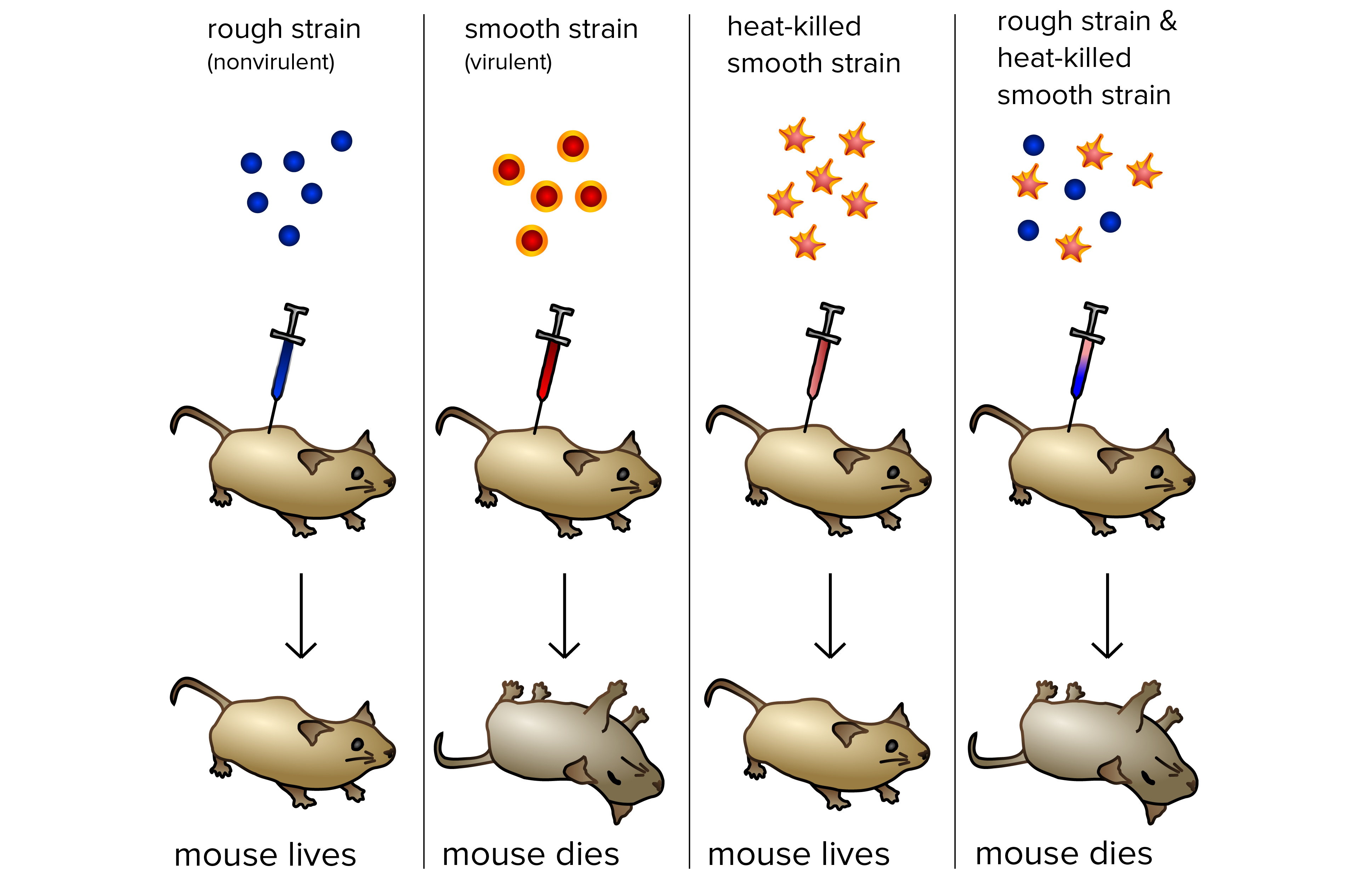
Frederick Griffith: transformação bacteriana
Em 1928, o bacteriologista britânico Frederick Griffith conduziu uma série de experimentos usando a bactéria Streptococcus pneumoniae e ratos. Griffith não estava tentando identificar o material genético, mas, sim, criar uma vacina contra a pneumonia. Em seus experimentos, Griffith usou duas cepas relacionadas de bactéria, conhecidas como R e S.
Cepa R. Quando cultivadas em placa de Petri, a bactéria R forma colônias ou aglomerados de bactérias relacionadas, que têm bordas bem definidas e aparência rugosa (daí a sigla "R"). As bactérias R são avirulentas, significando que não causam doenças quando injetadas em ratos.
Cepa S A bactéria S formou colônias arredondadas e suaves (daí a sigla "S"). A aparência suave se dá por conta de um polissacarídeo ou uma capa a base de açúcar produzida pela bactéria. Essa capa protegeu a bactéria S do sistema imune do rato, fazendo-as virulentos (capazes de causar doenças). Ratos em que foram injetadas bactérias S vivas desenvolveram pneumonia e morreram.
Como parte de seus experimentos, Griffith tentou injetar ratos com bactérias S mortas por calor (isso é, bactérias que haviam sido aquecidas a altas temperaturas, causando a morte das células). Sem surpresas, a bactéria S morta pelo calor não causou doença nos ratos.
Os experimentos tiveram um resultado inesperado, contudo, quando bactérias R inofensivas foram combinadas com bactérias S mortas por calor e injetadas em um rato. Não só o rato contraiu pneumonia e morreu, mas quando Griffith retirou uma amostra de sangue do rato morto, ele encontrou bactérias S vivas!
Griffith concluiu que as bactérias da cepa R teriam adquirido o que ele chamou de "princípio transformante" da bactéria S morta por calor, permitindo que elas se "transformassem" em bactérias S, tornando-se virulentas.
Avery, MacLeod e McCarty: Identificando o princípio transformante
Em 1944, três pesquisadores, canadenses e americanos, Oswald Avery, Maclyn McCarty, e Colin MacLeod, dispuseram-se a identificar o "princípio transformante" de Griffith.
Para tanto, eles começaram com grandes culturas de células S inativadas por calor e, através de uma longa série de etapas bioquímicas (determinadas por experimentação cuidadosa), purificaram progressivamente o princípio transformante através de lavagens, separação, ou destruição enzimática dos outros componentes celulares. Por este método, eles foram capazes de obter pequenas quantidades de princípio transformante altamente purificado, que eles puderam então analisar através de outros testes para determinar sua identidade
2squared.
Várias linhas de evidência sugeriram a Avery e seus colegas que o princípio transformante poderia ser DNA
2squared:
A substância purificada apresentou resultados negativos em testes químicos conhecidos para detectar proteínas, mas um resultado fortemente positivo em um teste químico conhecido para detectar DNA.
A composição dos elementos do princípio transformante purificado assemelhava-se muito a DNA em suas proporções de nitrogênio e fósforo.
Enzimas que degradam proteínas e RNA tinha pouco efeito sobre o princípio transformante, mas enzimas capazes de degradar DNA eliminavam a atividade transformante.
Todos estes resultados apontavam para DNA como o provável princípio transformante. Contudo, Avery foi cauteloso na interpretação de seus resultados. Ele percebeu que era possível que alguma substância contaminante presente em pequenas quantidades, não DNA, fosse o verdadeiro princípio transformante
3cubed.
Por causa desta possibilidade, o debate acerca da função do DNA continuou até 1952, quando Alfred Hershey e Martha Chase usaram uma abordagem diferente para conclusivamente identificar o DNA como o material genético.
Os Experimentos de Hershey-Chase
Em seus agora lendários experimentos, Hershey e Chase estudaram bacteriófagos, ou vírus que atacam bactérias. Os fagos que usaram era simples partículas compostas de proteína e DNA, com as estruturas externas feitas de proteína e o núcleo interno consistindo de DNA.
Hershey e Chase sabiam que os fagos prendiam-se à superfície de uma célula bacteriana hospedeira e injetavam alguma substância (DNA ou proteína) no hospedeiro. Esta substância dava "instruções" que faziam a bactéria hospedeira iniciar a produção de muitos e muitos fagos - em outras palavras, era o material genético do fago. Antes do experimento, Hershey pensou que o material genético se provaria ser proteína
4start superscript, 4, end superscript.
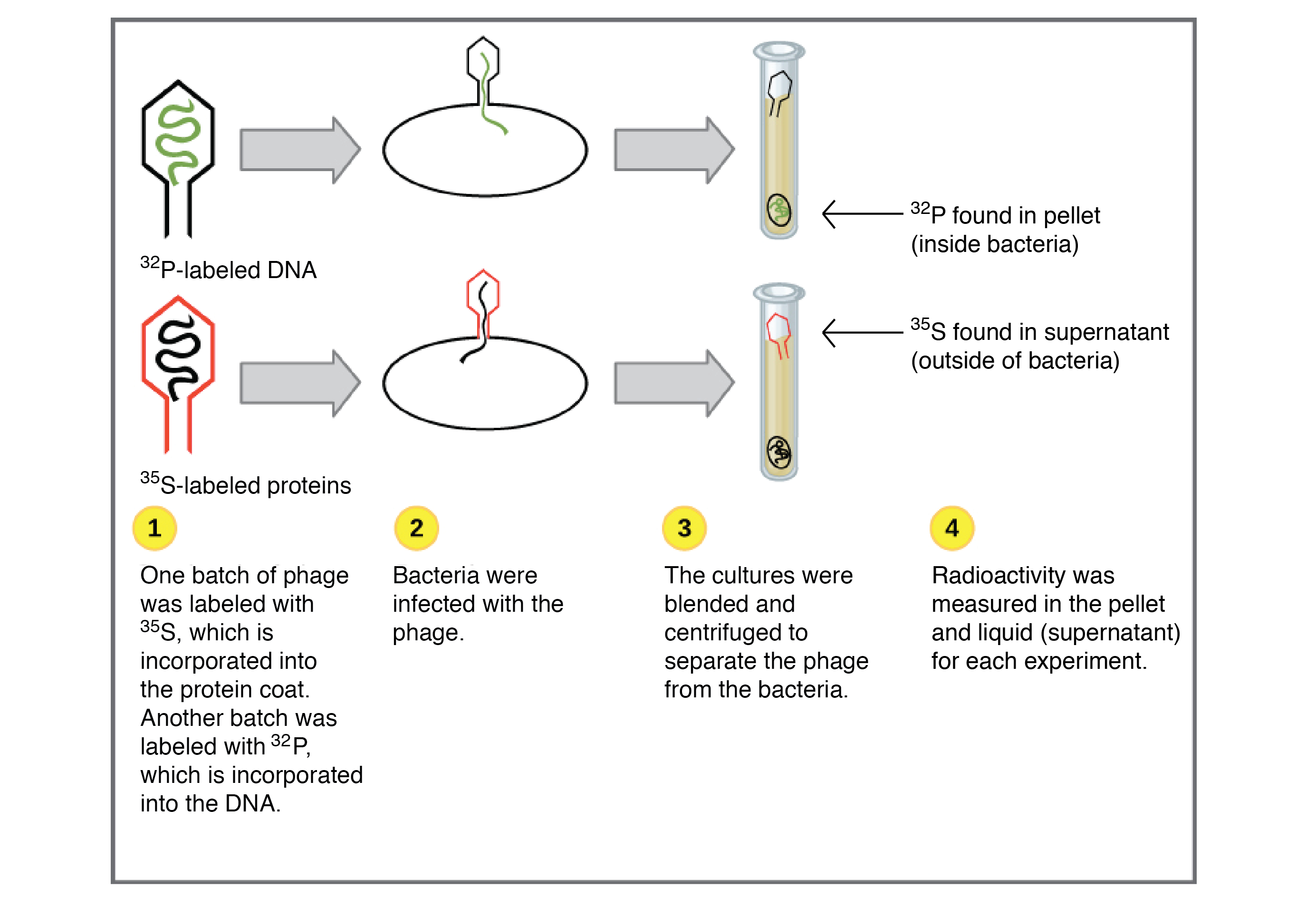
Para estabelecer se o fago injetava DNA ou proteína no interior da bactéria hospedeira, Hershey e Chase prepararam dois diferentes lotes de fagos. Em cada lote, os fagos eram produzidos na presença de elemento radiativo específico, que era incorporado nas macromoléculas (DNA e proteínas) sintetizadas pelos fagos.
Uma amostra foi produzida na presença de
35Sstart superscript, 35, end superscript, start text, S, end text, um isótopo radioativo do enxofre. O enxofre é encontrado em muitas proteínas e está ausente do DNA, assim somente as proteínas dos fagos eram radioativamente marcadas por esse tratamento.
A outra amostra foi produzida na presença de
32Pstart superscript, 32, end superscript, start text, P, end text, um isótopo radioativo de fósforo. O fósforo é encontrado no DNA mas não em proteínas, então só o DNA do fago (e não as proteínas do fago) estava marcado radioativamente por este procedimento.
Cada lote de fagos era usado para infectar uma cultura diferente de bactérias. Após a infecção, cada cultura era turbilhonada em um liquidificador, removendo qualquer fago remanescente e partes de fagos externas às células bacterianas. Finalmente, as culturas eram centrifugadas a alta velocidade, para separar as bactérias dos resíduos de fagos.
A centrifugação faz o material mais pesado, tais como bactérias, moverem-se para o fundo do tubo e formarem um amontoado chamado sedimentado. O material mais leve, tais como o meio (caldo) usado para o crescimento de culturas, junto com fagos e partes de fagos, permanecem próximo à parte de cima do tubo e forma uma camada líquida chamada de sobrenadante.
Quando Hershey e Chase mediram a radioatividade no sedimentado e no sobrenadante para ambos os experimentos, eles encontraram que uma grande quantidade de
32Pstart superscript, 32, end superscript, start text, P, end text apareceu no material sedimentado, enquanto quase todo o
35Sstart superscript, 35, end superscript, start text, S, end text apareceu no sobrenadante. Com base neste e em experimentos similares, Hershey e Chase concluíram que o DNA, não a proteína, era injetado nas células hospedeiras e compunham o material genético dos fagos.
Questões remanescentes
O trabalho dos pesquisadores acima providenciaram fortes evidencias de que o DNA é o material genético. No entanto, ainda não era claro como uma molécula supostamente simples poderia codificar a informação genética necessária para a construção de um organismo complexo. Pesquisas complementares de muitos cientistas, incluindo Erwin Chargaff, James Watson, Francis Crick e Rosalind Franklin levaram à
descoberta da estrutura do DNA, esclarecendo como o DNA pode codificar uma enorme quantidade de informações.
Créditos:
Referências:
Outras referências
Griffiths, A. J. F., Miller, J. H., Suzuki, D. T., et al. (2000). DNA: the genetic material. In
An introduction to genetic analysis (7th ed.). New York, NY: W. H. Freeman. Retirado de
http://www.ncbi.nlm.nih.gov/books/NBK22104/.
National Human Genome Research Institute. (2013, April 23). 1944: DNA is "transforming principle." No
Online education kit: 1940's. Retirado de
http://www.genome.gov/25520250.
Reece, J. B., Urry, L. A., Cain, M. L., Wasserman, S. A., Minorsky, P. V., and Jackson, R. B. (2011). DNA is the genetic material. Em Campbell biology (10th ed., pp. 313-315). San Francisco, CA: Pearson.
Exercícios de Aprendizagem:
1- (Enem 2005) Um fabricante afirma que um produto disponível comercialmente possui DNA vegetal, elemento que proporcionaria melhor hidratação dos cabelos.
Sobre as características químicas dessa molécula essencial à vida, é correto afirmar que o DNA
a) de qualquer espécie serviria, já que têm a mesma composição.
b) de origem vegetal é diferente quimicamente dos demais, pois possui clorofila.
c) das bactérias poderia causar mutações no couro cabeludo.
d) dos animais encontra-se sempre enovelado e é de difícil absorção.
e) de características básicas assegura sua eficiência hidratante.
2- (Ufrgs 2013) Sabe-se que a replicação do DNA é semiconservativa. Com base nesse mecanismo de replicação, assinale com V (verdadeiro) ou F (falso) as afirmações abaixo.
( ) O DNA original atua como molde, e cada novo DNA possui uma fita antiga e outra nova.
( ) Os quatro ribonucleosídeos trifosfatados, dATP, dGTP, dCTP e dUTP, devem estar presentes.
( ) O DNA deve ser desnaturado (desenrolado) para tornar-se acessível ao pareamento das novas bases.
( ) A enzima DNA polimerase adiciona nucleotídeos novos de acordo com o molde de DNA.
A sequência correta de preenchimento dos parênteses, de cima para baixo, é
a) V – V – F – F.
b) F – V – V – V.
c) V – F – V – V.
d) F – V – F – F.
e) F – F – F – V.
3- (Udesc 2016) Analise as proposições, em relação aos ácidos nucleicos, e assinale (V) para verdadeira e (F) para falsa.
( ) Os ácidos nucleicos são moléculas gigantes formadas por unidades chamadas de nucleotídeos.
( ) O RNA transportador é formado a partir de regiões específicas do DNA.
( ) O RNA ribossômico associado com proteínas forma os ribossomos.
( ) O DNA apresenta-se altamente condensado nas células procarióticas.
Assinale a alternativa que apresenta a sequência correta, de cima para baixo.
a) F – V – V – F
b) F – F – V – V
c) V – V – V – F
d) V – F – V – V
e) V – F – F – V