2° Ano E Biologia Profa Solange
Período para entrega: Até 16/09/2020
Unidade: DNA como material genético- DNA- Receita da Vida e seu Código.
Instruções:
1- Fazer a Leitura e Interpretação de Texto- (Khan Academy)- Texto 1- Descoberta da estrutura do DNA
Texto 2- Experimentos Clássicos: DNA como material genético
2- Responder as questões disponibilizadas abaixo dos textos.
Descoberta da estrutura do DNA
Introdução

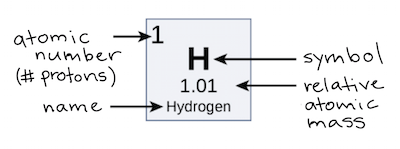
Os componentes do DNA
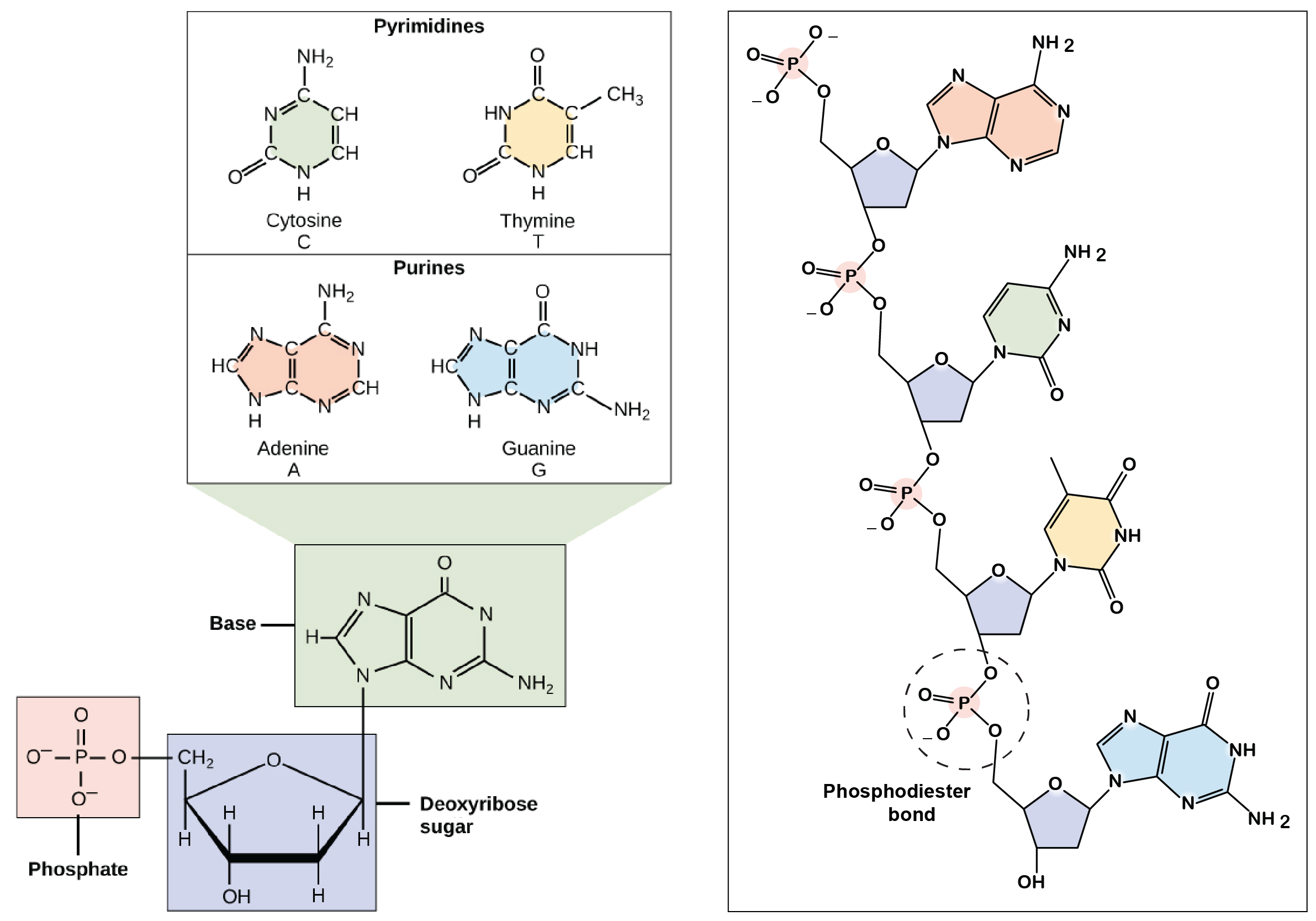
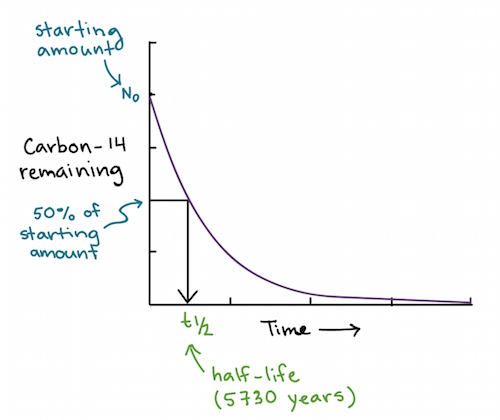
Regras de Chargaff
- A, T, C e G não eram encontradas em quantidades iguais (como alguns modelos da época diziam)
- As quantidades de bases variavam entre as espécies, mas não entre indivíduos da mesma espécie
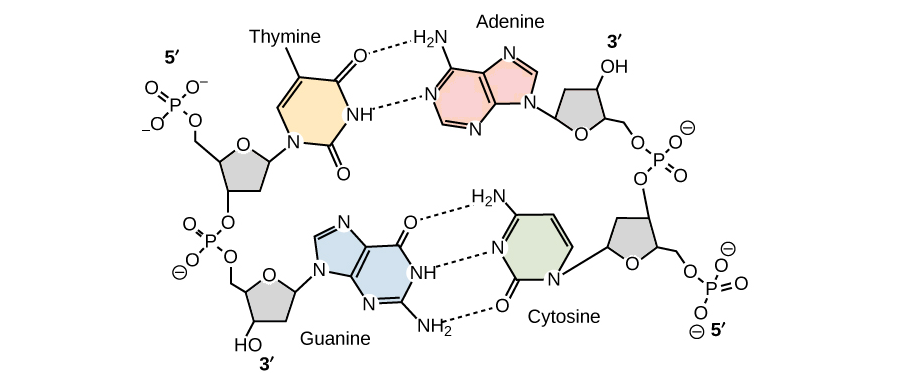
- A quantidade de A sempre igualava a quantidade de T, e a quantidade de C sempre igualava a quantidade de G (A = T e G = C)
Watson, Crick e Rosalind Franklin

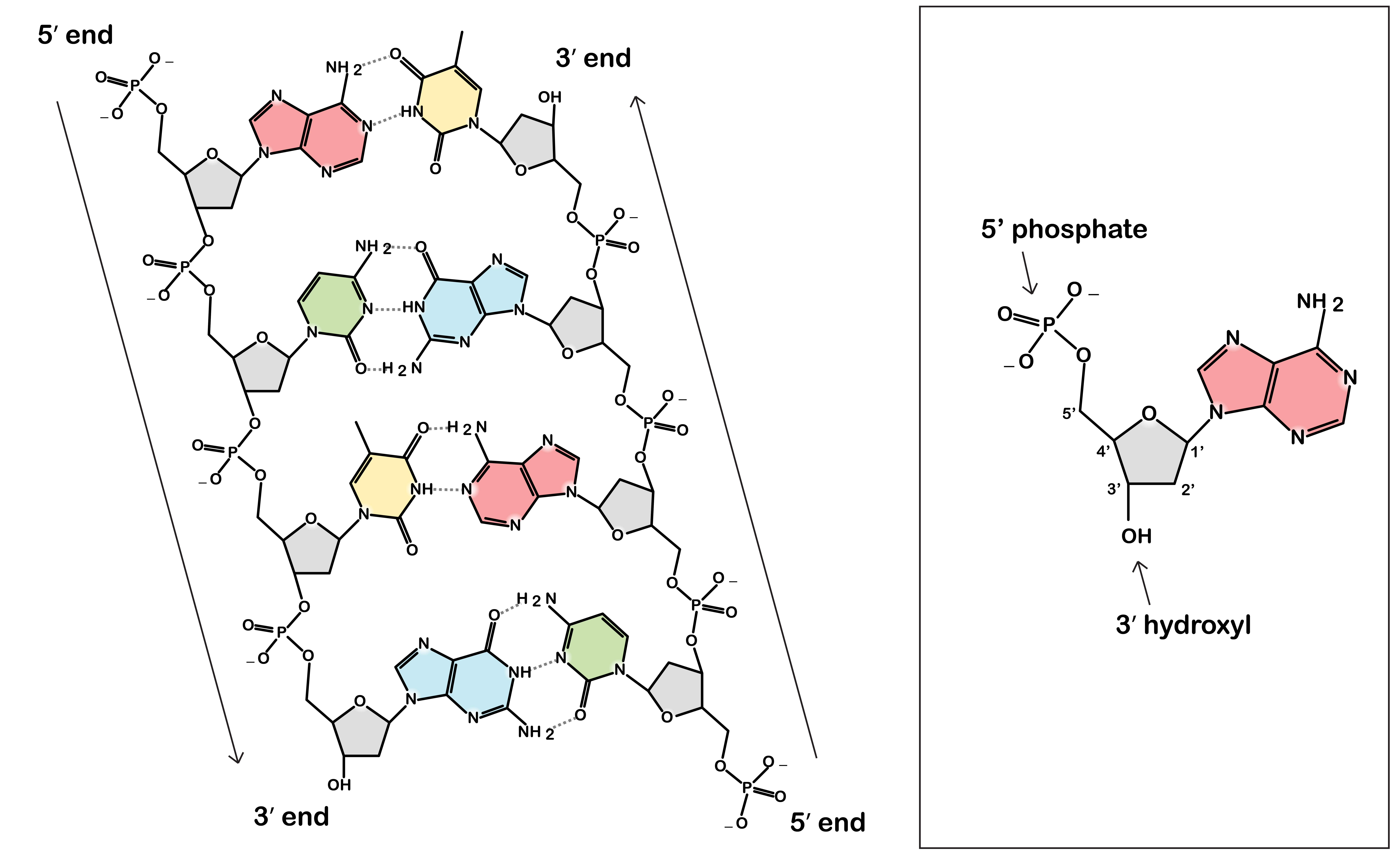
O modelo do DNA de Watson e Crick
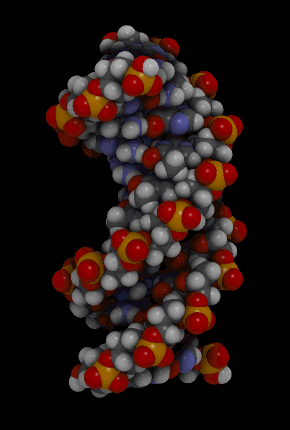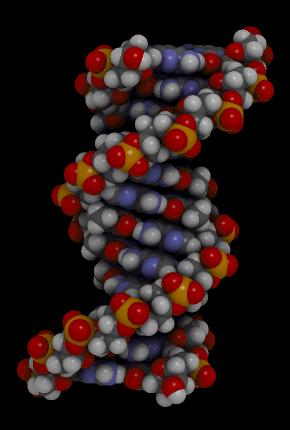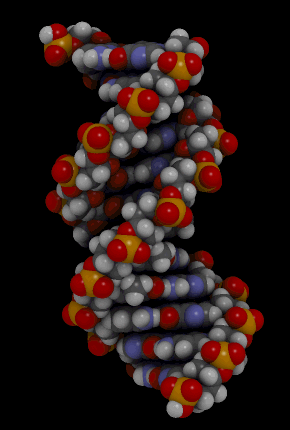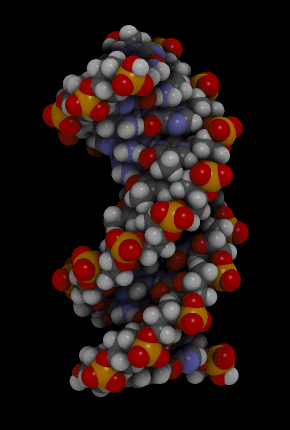
Orientação antiparalela
Hélice dextrógira
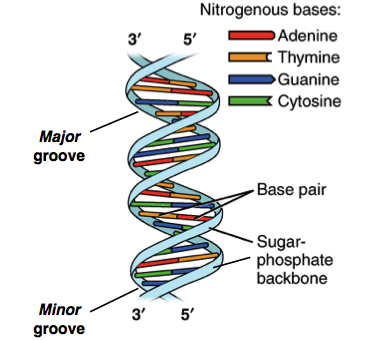
Pareamento de bases
O impacto da dupla hélice
Explore além da Khan Academy
Créditos:
- "DNA structure and sequencing," by OpenStax College, BIology, CC BY 4.0. Baixe o artigo original gratuitamente em http://cnx.org/contents/185cbf87-c72e-48f5-b51e-f14f21b5eabd@10.53.
- "Historical basis of modern understanding," by OpenStax College, Biology, CC BY 4.0. Baixe o artigo original gratuitamente em http://cnx.org/contents/185cbf87-c72e-48f5-b51e-f14f21b5eabd@10.53.
Referências:
- Pray, L. A. (2008). Discovery of DNA structure and function: Watson and Crick. Nature Education, 1(1), 100. Disponível em http://www.nature.com/scitable/topicpage/discovery-of-dna-structure-and-function-watson-397.
- Aldridge, S. The DNA story. In Royal society of chemistry. 2003. Disponível em http://www.rsc.org/chemistryworld/Issues/2003/April/story.asp. Acesso em 27 jul 2016.
- Cobb, M. (2015, June 23). Sexism in science: Did Watson and Crick really steal Rosalind Franklin's data? The Guardian. Disponível em http://www.theguardian.com/science/2015/jun/23/sexism-in-science-did-watson-and-crick-really-steal-rosalind-franklins-data.
- The DNA riddle: King's College, London, 1951-1953. (n.d.) In The Rosalind Franklin papers. Disponível em https://profiles.nlm.nih.gov/ps/retrieve/Narrative/KR/p-nid/187.
- Dugard, J. (2003, March 18). A grave injustice. Mail & Guardian. Disponível em http://mg.co.za/article/2003-03-18-a-grave-injustice.
- Tyson, P. (2003, April 22). Rosalind Franklin's legacy. In NOVA. Disponível em http://www.pbs.org/wgbh/nova/tech/rosalind-franklin-legacy.html.
- Rosalind Franklin. (2016, January 15). Acesso em 15 de janeiro, 2016. Disponível em Wikipedia: https://en.wikipedia.org/wiki/Rosalind_Franklin.
- Banquet speech: James Watson's speech at the Nobel banquet in Stockholm, December 10, 1962. (2016). In Nobelprize.org. Disponível em http://www.nobelprize.org/nobel_prizes/medicine/laureates/1962/watson-speech.html.
- Molecular structure and function: Evolution with a twist. (n.d.) In Biology-101: Brantley. Disponível em http://www.science-projects.com/Helices.htm. Acesso em 27 de julho, 2016.
- B-form, A-form, and Z-form of DNA. (2014, May 4). Acesso em 27 de julho, 2016. Disponível em bio.libretexts.org/Core/Genetics/Unit_I%3A_Genes,_Nucleic_Acids,_Genomes_and_Chromosomes/2%3A_Structures_of_nucleic_acids/B-Form,_A-Form,_Z-Form_of_DNA.
- Cambridge Physics. (n.d). A working model! In The structure of DNA: Crick and Watson, 1953. Disponível em http://www-outreach.phy.cam.ac.uk/camphy/dna/dna14_1.htm.
- Watson, J. D; Crick, F. H. C. A structure for deoxyribose nucleic acid. Nature, v. 171, n. 4356, pp. 737-738. Disponível em http://www.nature.com/nature/dna50/watsoncrick.pdf.
Referências:
Experimentos Clássicos: DNA como material genético
Introdução
Proteína x DNA
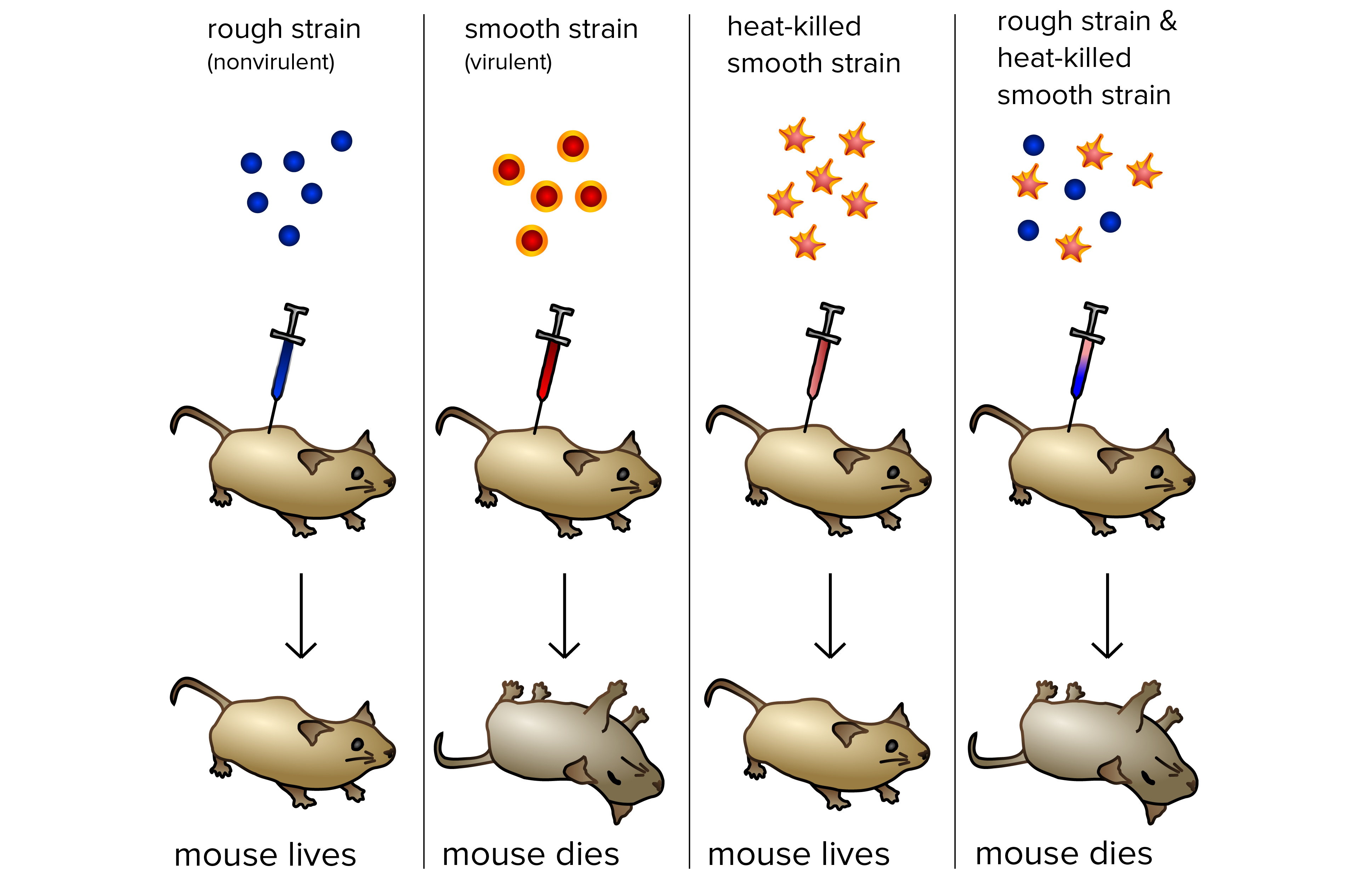
Frederick Griffith: transformação bacteriana
- Cepa R. Quando cultivadas em placa de Petri, a bactéria R forma colônias ou aglomerados de bactérias relacionadas, que têm bordas bem definidas e aparência rugosa (daí a sigla "R"). As bactérias R são avirulentas, significando que não causam doenças quando injetadas em ratos.
- Cepa S A bactéria S formou colônias arredondadas e suaves (daí a sigla "S"). A aparência suave se dá por conta de um polissacarídeo ou uma capa a base de açúcar produzida pela bactéria. Essa capa protegeu a bactéria S do sistema imune do rato, fazendo-as virulentos (capazes de causar doenças). Ratos em que foram injetadas bactérias S vivas desenvolveram pneumonia e morreram.
Avery, MacLeod e McCarty: Identificando o princípio transformante
- A substância purificada apresentou resultados negativos em testes químicos conhecidos para detectar proteínas, mas um resultado fortemente positivo em um teste químico conhecido para detectar DNA.
- A composição dos elementos do princípio transformante purificado assemelhava-se muito a DNA em suas proporções de nitrogênio e fósforo.
- Enzimas que degradam proteínas e RNA tinha pouco efeito sobre o princípio transformante, mas enzimas capazes de degradar DNA eliminavam a atividade transformante.
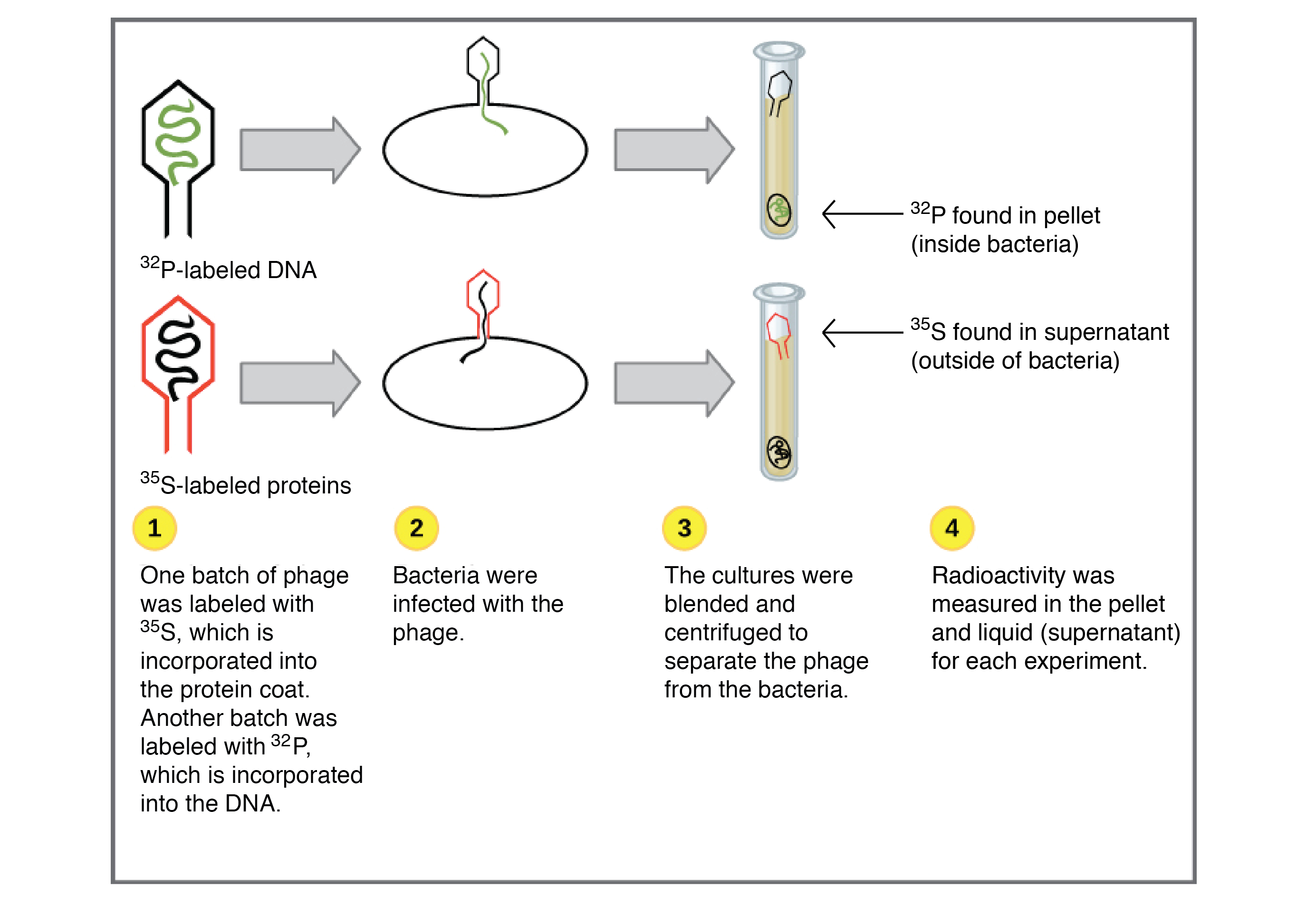
Os Experimentos de Hershey-Chase
- Uma amostra foi produzida na presença de start superscript, 35, end superscript, start text, S, end text, um isótopo radioativo do enxofre. O enxofre é encontrado em muitas proteínas e está ausente do DNA, assim somente as proteínas dos fagos eram radioativamente marcadas por esse tratamento.
- A outra amostra foi produzida na presença de start superscript, 32, end superscript, start text, P, end text, um isótopo radioativo de fósforo. O fósforo é encontrado no DNA mas não em proteínas, então só o DNA do fago (e não as proteínas do fago) estava marcado radioativamente por este procedimento.
Questões remanescentes
Créditos:
Referências:
- Aldridge, Susan. (2003). The DNA story. In Royal society of chemistry. Disponível em: http://www.rsc.org/chemistryworld/Issues/2003/April/story.asp. Acesso em 27 jul. 2016.
- Avery, O. T., MacLeod, C. M., and McCarty, M. (1944). Studies on the chemical nature of the substance inducing transformation of Pneumococcal types. J. Exp. Med., 79(2), 137-158. Disponível em: http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2135445/.
- Scarc. (2009, July 7). Oswald Avery's Pneumococcus experiments: Forerunner of the DNA story [web log post]. In The Pauling blog. Disponível em: https://paulingblog.wordpress.com/2009/07/07/oswald-averys-pneumococcus-experiments-forerunner-of-the-dna-story/.
- Scarc. (2009, August 18). The Hershey-Chase blender experiments [web log post]. In The Pauling blog. Retrieved from https://paulingblog.wordpress.com/2009/08/18/the-hershey-chase-blender-experiments/.
Outras referências
Sobre as características químicas dessa molécula essencial à vida, é correto afirmar que o DNA
a) de qualquer espécie serviria, já que têm a mesma composição.
b) de origem vegetal é diferente quimicamente dos demais, pois possui clorofila.
c) das bactérias poderia causar mutações no couro cabeludo.
d) dos animais encontra-se sempre enovelado e é de difícil absorção.
e) de características básicas assegura sua eficiência hidratante.
2- (Ufrgs 2013) Sabe-se que a replicação do DNA é semiconservativa. Com base nesse mecanismo de replicação, assinale com V (verdadeiro) ou F (falso) as afirmações abaixo.
( ) O DNA original atua como molde, e cada novo DNA possui uma fita antiga e outra nova.
( ) Os quatro ribonucleosídeos trifosfatados, dATP, dGTP, dCTP e dUTP, devem estar presentes.
( ) O DNA deve ser desnaturado (desenrolado) para tornar-se acessível ao pareamento das novas bases.
( ) A enzima DNA polimerase adiciona nucleotídeos novos de acordo com o molde de DNA.
A sequência correta de preenchimento dos parênteses, de cima para baixo, é
a) V – V – F – F.
b) F – V – V – V.
c) V – F – V – V.
d) F – V – F – F.
e) F – F – F – V.
3- (Udesc 2016) Analise as proposições, em relação aos ácidos nucleicos, e assinale (V) para verdadeira e (F) para falsa.
( ) Os ácidos nucleicos são moléculas gigantes formadas por unidades chamadas de nucleotídeos.
( ) O RNA transportador é formado a partir de regiões específicas do DNA.
( ) O RNA ribossômico associado com proteínas forma os ribossomos.
( ) O DNA apresenta-se altamente condensado nas células procarióticas.
Assinale a alternativa que apresenta a sequência correta, de cima para baixo.
a) F – V – V – F
b) F – F – V – V
c) V – V – V – F
d) V – F – V – V
e) V – F – F – V